WORKING GROUP REPORTS
Latitudinal Gradients Within the Eastern Boundary Current System
Cochairs: M. Ohman and B. Hickey
Participants: V. Holliday, A. Huyer, P. Kremer, A. MacCall, D. Mackas, K. Parker, W. Pearcy, J.
Schumacher, T. Strub, R. Tipper, and D. Ware
INTRODUCTION
The California Current system (CCS) comprises three broadly definable regions: I, Vancouver Island
to Cape Blanco; II, Cape Blanco to Point Conception; and III, Point Conception to northern Baja
California (Fig. 5). These regions are characterized by differences in wind stress, intensity of coastal
upwelling, coastal morphology, freshwater inflow, and influence of long-time-scale advection. The
regions also differ in seasonality of planktonic production cycles. For some planktonic species, the
regional boundaries represent biogeographic boundaries, although other species range throughout the
three regions.
Among the most striking biological contrasts is the paucity of surface spawning by epipelagic fishes in
the central region (II; see Parrish et al. 1981). This region is characterized by strong seasonal
upwelling near coastal promontories and by a variety of mesoscale features such as coastal jets,
eddies, and filaments that tend to transport organisms offshore. The fish species that do reproduce in
this region tend to brood eggs or larvae, or to spawn demersally in protected waters. Equally striking
is the concentration of spawning activity in Region III: about 90% of the epipelagic fish biomass
(hake, sardine, anchovy) in the southern part of the California Current system spawns in the Southern
California Bight and waters offshore. Primary and secondary production in Region III is therefore
important to a significant fraction of California Current fishes.
A number of critical physical processes are likely to govern much of the variability in marine
populations over time in the CCS and other eastern boundary currents:
- Mesoscale eddies, jets, and meanders
- Timing, duration, and intensity of coastal upwelling
- Vertical mixing events
- Alongshore advection
These processes may be influenced by future changes in the global ocean; some evidence suggests that
changes have already occurred in recent decades. The differences in physical-biological linkages in
Regions I, II, and III provide a natural basis for comparative study of the changes in marine
populations that may accompany different scenarios for climate change.
We strongly recommend conducting comparative studies in all three regions of the CCS. Forcing
functions differ spatially, but some marine species depend on processes in all three regions, so a
successful GLOBEC program will have to incorporate the significant processes operating on a broad
latitudinal scale. One component of this program should include intensive field studies that focus on
processes thought to be important in each region. As a second component, we recommend time series
sampling in each of the three regions for at least a decade. The time series might
include a combination of moored arrays, satellite observations, and ship surveys. Suitable localities
within each region might include the west coast of Vancouver Island, or the coasts of Washington or
northern Oregon (Region I); Point Arena to Point Sur (Region II); and the Southern California Bight
(Region III). In each region there are oceanographic institutions and shipboard resources that could be
applied to the task. It is likely that existing field programs in each of the three regions could be
expanded.
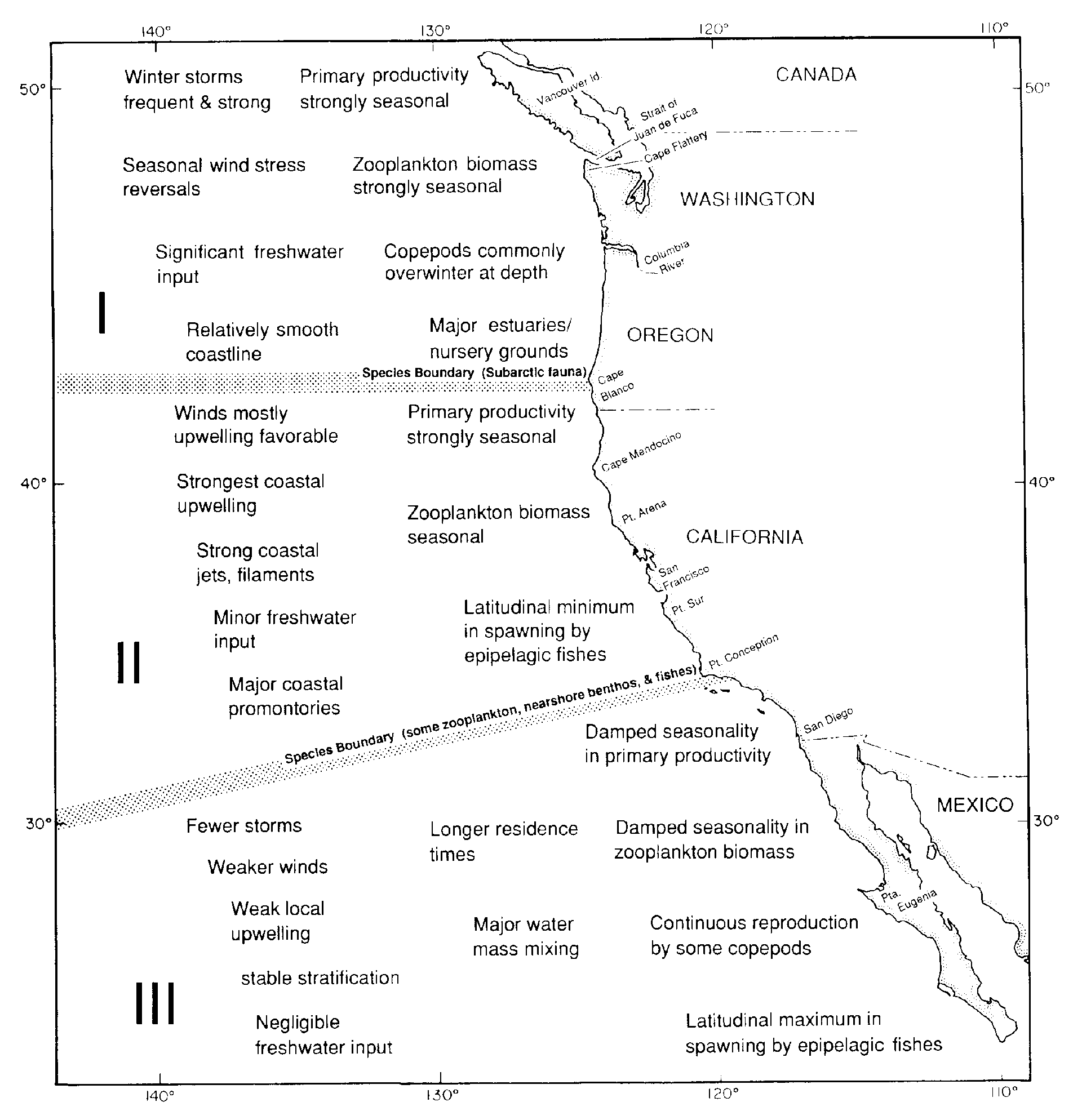
Figure 5. Generalized regional variations in physical and biological
processes within the California Current System. The boundaries
between Regions I, II, and III are only approximate and vary over
time. The generalizations regarding Region III apply primarily to
the Southern California Bight.
REGIONS OF THE CALIFORNIA CURRENT SYSTEM
The California Current system spans more than 20 degrees of latitude along the west coast of North
America. Its range exceeds the scales of dominant atmospheric pressure systems as well as the scales
of regional coastal morphology. Thus the variability of the California Current and associated
planktonic populations inherently includes significant and sometimes dramatic latitudinal differences.
Many of these differences (e.g., wind forcing, temperature, photoperiod) are roughly a linear function
of latitude, whereas others have distinct biogeographical boundaries. Examination of the dominant
characteristics suggests that the California Current has three principal regions (Fig. 5). The boundaries
between these regions are not exact, particularly between Regions I and II, where the area of
seasonally reversing monthly mean winds extends farther south (to about 39°N) than the area of
relatively straight coastline (about 42°N). The boundaries are known to change during El
Niño/Southern Oscillation and other long-term events, including intrusions of subarctic water from the
north. The boundaries might also be expected to shift under some scenarios of global change.
In Region I, coastal wind stress is relatively strong, and wind direction reverses seasonally as well as
on shorter time scales. Winter storms are particularly strong and frequent, leading to intense mixing
and alongshore northward advection (Huyer et al. 1978; Thomson 1981; Hickey 1989; Thomson et al.
1989). Except for the region near the Strait of Juan de Fuca, the coastline is relatively straight, and the
shelf is continuous, though narrow, over large alongshore scales (hundreds of km). Significant
freshwater input is provided throughout most of the year by the Strait of Juan de Fuca and by the
Columbia River. Large estuaries are relatively common and are thought to provide nurseries for
several important species (e.g., Dungeness crab, McConnaughey et al., in press; Pacific herring,
Haegele and Schweigert 1985). Primary production rates (Perry et al. 1989) and zooplankton biomass
(Mackas 1992) have strong seasonal variations in this region. Some of the major copepod species
overwinter at depth, then reappear in the surface layer for relatively short periods of growth in spring
and summer. Several species (e.g., Neocalanus plumchrus, N. cristatus, Eucalanus bungii, Calanus
pacificus oceanicus) enter Region I from the Subarctic Pacific or the West Wind Drift (Fleminger
1964; Fleminger and Hulseman 1973) and rarely extend south of this region.
The dominant physical characteristics of Region II, which extends approximately from Cape Blanco to
Point Conception, are the coastal promontories. Recent research suggests that energetic coastal jets,
filaments, and meanders are associated with these promontories (Davis 1985; Kosro & Huyer 1986;
Huyer & Kosro 1987; Strub et al. 1991). Current jets commonly extend 200-300 km offshore and
may lead to relatively short residence times for plankton in the coastal zone. The strongest equatorward
wind stress and, hence, coastal upwelling also occur in Region II (Nelson 1977; Huyer 1983; Strub et
al. 1987). Although wind stress varies seasonally, the seasonal mean is always directed toward the
equator. The strong coastal upwelling in this region supplies "new" nutrients into the euphotic zone,
leading to elevated primary production rates and high standing stocks of phytoplankton (Dugdale &
Wilkerson 1989). Satellite imagery suggests that many of the high-chlorophyll features found in
Region II are associated with jets, eddies, and other mesoscale features (Flament et al. 1985).
Zooplankton biomass varies seasonally (Roesler & Chelton 1987). Zooplankton species composition
can shift relatively abruptly at the frontal boundaries associated with mesoscale jets and eddies. Among
the most dramatic biological characteristics of Region II is the latitudinal minimum in spawning of
pelagic fishes. Whereas epipelagic fishes spawn extensively in Region III and to some extent in
Region I, those in Region II appear mainly to brood their eggs or larvae (e.g., rockfishes) or to use
nearshore embayments as spawning and nursery grounds (e.g., Pacific herring) that appear to reduce
the probability of offshore transport of pelagic larvae (Parrish et al. 1981).
The dominant physical characteristic of Region III is that, because of the coastline bend at Point
Conception, local wind stress is relatively weak on the scales of seasons and events (Nelson 1977;
Halliwell and Allen 1987). Thus local upwelling is weak in spring and summer, and wind- and
wave-induced mixing is relatively weak year-round. Winter storms occur only occasionally.
Freshwater input is insignificant. Interleaving of differing water masses occurs in Region III, making
it particularly sensitive to large-scale, long-time-scale environmental perturbations such as ENSO
(Hickey 1979; Lynn & Simpson 1987,1990; Tsuchiya 1980). Seasonal cycles in zooplankton biomass
are relatively weak (Roesler & Chelton 1987). Deep overwintering of calanoid copepods occurs
(Alldredge et al. 1984), but it may involve only part of a population while another part grows and
reproduces year-round (Mullin & Brooks 1967). The boundary between Region II and III is a
biogeographic boundary for some species of nearshore benthic marine invertebrates and pelagic fishes.
Region III is the preferred spawning site for over 90% of the epipelagic fish biomass (hake, sardine,
anchovy) in the southern part of the CCS.
In addition to these latitudinal patterns, strong cross-shore variations occur in the CCS. For example,
the wind field has strong cross-shore gradients at most latitudes, with maximum winds occurring
seaward of the continental shelf (Nelson 1977). Vertically integrated primary production rates tend to
decrease in the cross-shore direction (P.E. Smith, pers. comm.; F. Chavez pers. comm; Perry et al.
1989). A zone of maximum variability in dynamic height begins approximately 200-300 km offshore
(Lynn & Simpson 1987); this zone has been called an eddy alley. The long-term maximum in
macrozooplankton biomass occurs offshore in some areas (Roesler & Chelton 1987). This maximum
is sometimes dominated by gelatinous zooplankton such as salps and doliolids (Berner 1967).
CRITICAL EASTERN BOUNDARY CURRENT PROCESSES AND BIOLOGICAL
LINKAGES
We identify key physical-biological linkages that should be addressed in the GLOBEC eastern
boundary current program. Each linkage focuses on physical processes thought to significantly affect
population growth rates of metazoans having a planktonic stage. The relative importance of these
processes typically differs in the three regions of the CCS. The regional differences may be viewed as
a "natural experiment," permitting the relative impact of different processes to be quantified and
perhaps used as a basis for projecting responses of marine populations to different scenarios of global
change. We also pose a few preliminary hypotheses; we expect these to be modified and others to be
put forward in future discussions.
Mesoscale Eddies, Jets, and Meanders
Rationale: Physical features such as eddies, jets, and meanders of coastal currents can be highly
energetic and variable over time. Planktonic organisms entrained within these features often experience
increased offshore (or onshore) transport, as well as different regimes of food and predators.
Behavioral adaptations to such features (e.g., diel vertical migration interacting with current shear) can
increase the residence time of organisms in the coastal zone.
H1: Nearshore eddies, jets, and current meanders are significant dispersal mechanisms for coastal
populations.
H2: Frontal zones associated with these mesoscale features are sites of enhanced production and
concentration of planktonic prey.
H3: Offshore mesoscale eddies are retention sites that reduce spatial losses and enhance population
growth rates of some planktonic populations.
Timing, Duration, and Intensity of Coastal Upwelling
Rationale: Coastal upwelling influences planktonic organisms through a number of mechanisms.
These include: (1) food-web effects (mediated by the input of "new" nutrients and elevated primary
production); (2) seeding of epipelagic populations from dormant stages in deeper water; and (3)
offshore transport of organisms entrained in upwelling filaments and jets. We emphasize the first two
mechanisms here.
H1: Seeding from dormant stages is more important to population growth in Region II, where
offshore transport is more frequent, than in Region I, where transport is more frequently alongshore.
H2: A significant fraction of the primary and secondary production in Region II is advected off the
shelf and is unavailable to pelagic fishes that normally inhabit the continental shelf and slope (e.g.,
Pacific hake and northern anchovy).
Vertical Mixing Events
Rationale: (1) The shape and depth of the pycnocline affects the supply of new nutrients into the
euphotic zone. This alters the magnitude of primary production as well as food-web structure (i.e., the
size and species composition of phytoplankton and microzooplankton). Vertical mixing can change the
pycnocline topography and thus food-web structure. (2) Lasker's "stable ocean hypothesis" suggests
that "turbulent" mixing can erode the microscale aggregations of microplankton that are essential for
the first-feeding success of some larval fishes. The intensity and frequency of storms can alter the
availability of prey to a variety of different zooplankters. (3) Turbulence can affect the encounter rates
of prey and predators. Recent studies suggest that encounter rates increase with turbulent kinetic
energy dissipation (Yamazaki et al. 1991).
H1: There is an optimal wind speed that maximizes primary and hence secondary production. The
optimal speed is about 7-8 m s-l.
H2: Survival is greatest for fish larvae that hatch during calm conditions.
Alongshore Advection: Long Time Scales
Rationale: Biogeographic boundaries exist within the CCS, e.g., at Point Conception (the boundary of
Regions II and III) and sometimes at the boundary between Regions I and II (Brinton 1962). With
alterations in the large-scale circulation of the CCS, these biogeographic boundaries may shift, thereby
affecting the survival patterns and spawning success of indigenous populations. Circulation changes
may also introduce faunal elements from different biogeographic provinces (e.g., the subarctic Pacific
or tropical waters; Pearcy and Schoener 1987). Benthic populations dependent upon particular
substrates or sediment characteristics may change abruptly and nonlinearly if the required substrates
are discontinuous along the coast and animals are displaced considerable distances alongshore.
H1: Large-scale advection from the north alters the species composition and secondary production of
CCS zooplankton assemblages.
H2: Large-scale advection from the south during El Niño/Southern Oscillation alters the species
composition and secondary production of CCS zooplankton assemblages.
Alongshore Advection: Shorter Time Scales
Rationale: "Spatial losses" of organisms from a desirable habitat or spawning grounds can result in
marked depression of recruitment success (e.g., Bailey et al. 1982). Conversely, alongshore advection
can also serve as an essential mechanism by allowing planktonic organisms to return upstream to
favorable spawning grounds or habitats. Some aspects of alongshore advection are relatively
predictable (e.g., seasonal reversal of winds and currents in Region I; the seasonal in Region I but not
in Region III.
H1: Some pelagic species need the poleward-flowing California Undercurrent and Davidson Current
to complete their life history.
Each of these processes may fluctuate on a variety of time scales. For example, the frequency or
intensity of upwelling-favorable winds may change within a single season, as well as over decades
(Bakun 1990).
RESEARCH STRATEGIES
We recommend that two research strategies be pursued in parallel. The first is to study a long-lived,
dominant species throughout its life history as it migrates through the different regions of the CCS.
This might be characterized as a Lagrangian mode of study. The study site will shift with the ontogeny
of the organism to investigate the processes leading to spawning success, larval and juvenile survival,
latitudinal migrations, and adult feeding success. A particularly good candidate for this approach is the
Pacific hake. Hake spawn in a geographically restricted area in the west of Region III, yet eventually
migrate northward to Region I, where they are found as adults (Fig. 6). They may be seen as
"integrators" of different processes in different regions of the CCS and therefore particularly sensitive
to changes in ocean circulation and food-web structure. Preliminary evidence suggests that their
spawning grounds moved northward in the late 1970s (inset, Fig. 6). The timing of this shift
corresponds to a warming period in the ocean (A. MacCall, pers. comm.). Thus Pacific hake may be
particularly sensitive to global change.
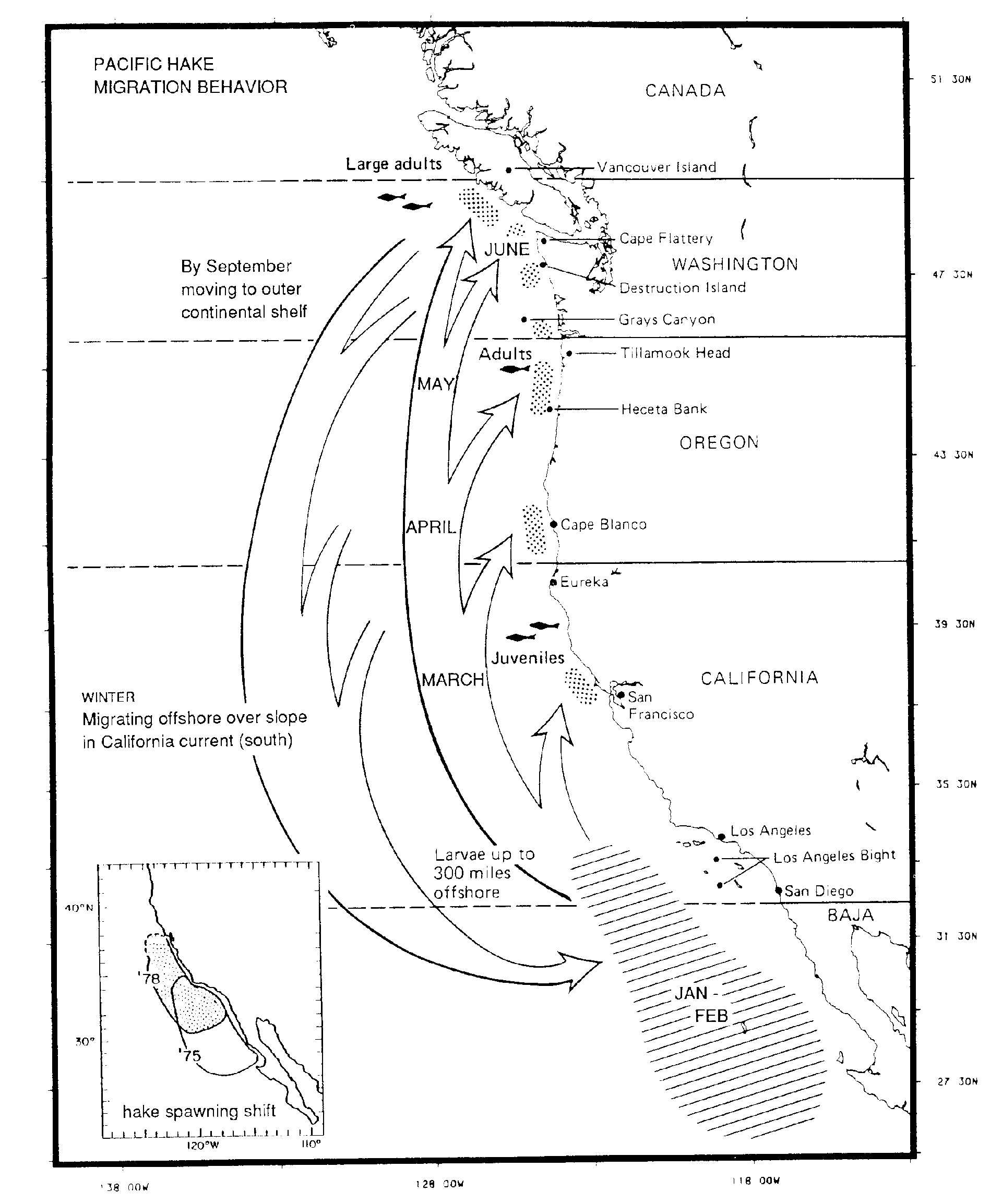
Figure 6. Migration of the Pacific hake, Merluccius productus,
from Bailey et al. 1982; interpretation of timing modified by D. M.
Ware. Inset illustrates the geographic shift in spawning area from
12975 to 1978 (P. E. Smith, pers. comm.). The northward displacement
of spawing occurred at a time of warming in the California Current
system (A. D. MacCall, pers. comm.).
The second research strategy involves selecting species (or sibling species) that occur broadly
throughout the CCS. It is hypothesized that the same species are governed by different processes in
different regions. This strategy might be viewed as an Eulerian mode of study. In the different regions
occupied by "metapopulations," or subpopulations, of the species, the differing effects of processes
such as offshore transport, food limitation, vertical mixing, or large-scale advection can be quantified.
A simplified description of the population growth rate for a metapopulation within each of the three
regions of the CCS can be expressed as follows:
dNR / dt = birthR - deathR - horizontal migrationR - advectionR + diffusionR
where the subscript R designates the region of interest. The terms in the equation are each rather
complex and nonlinear functions of other processes and will vary within (as well as between) regions.
Nevertheless, in this Eulerian approach, strong regional contrasts in the importance of these terms
should make it possible to identify the most significant population control mechanisms. For example,
for cyprid larvae of barnacles, the advection term may predominate in Region II, and the death rate
term may predominate in Region I.
TARGET SPECIES
Fish
Approximately 90% of the epipelagic fish biomass in the southern part of the California Current
spawns in the southern California Bight and associated offshore waters (D. Ware, pers. comm.). The
three primary species are Pacific hake, northern anchovy, and sardine (Ware and McFarlane 1989).
Because hake constitute 50-60% of all epipelagic fish biomass, they are a particularly suitable target
for this study, in addition to the reasons identified above. Recent evidence for recovery of the sardine
(P. Smith, pers. comm.), combined with extensive historical information on the sardine and northern
anchovy, suggests that they are also excellent candidates for study. Pacific hake and anchovy have
spawning subpopulations in Region I as well as Region III and thus are good candidates for the
Eulerian approach.
Although the spawning regions of the Pacific hake and northern anchovy overlap broadly, the two
species show markedly different interannual variations in recruitment (Fig. 7). Hake have occasional
very strong year classes, while northern anchovy tend to have runs of weaker or stronger year classes.
This difference is a likely topic for GLOBEC studies.
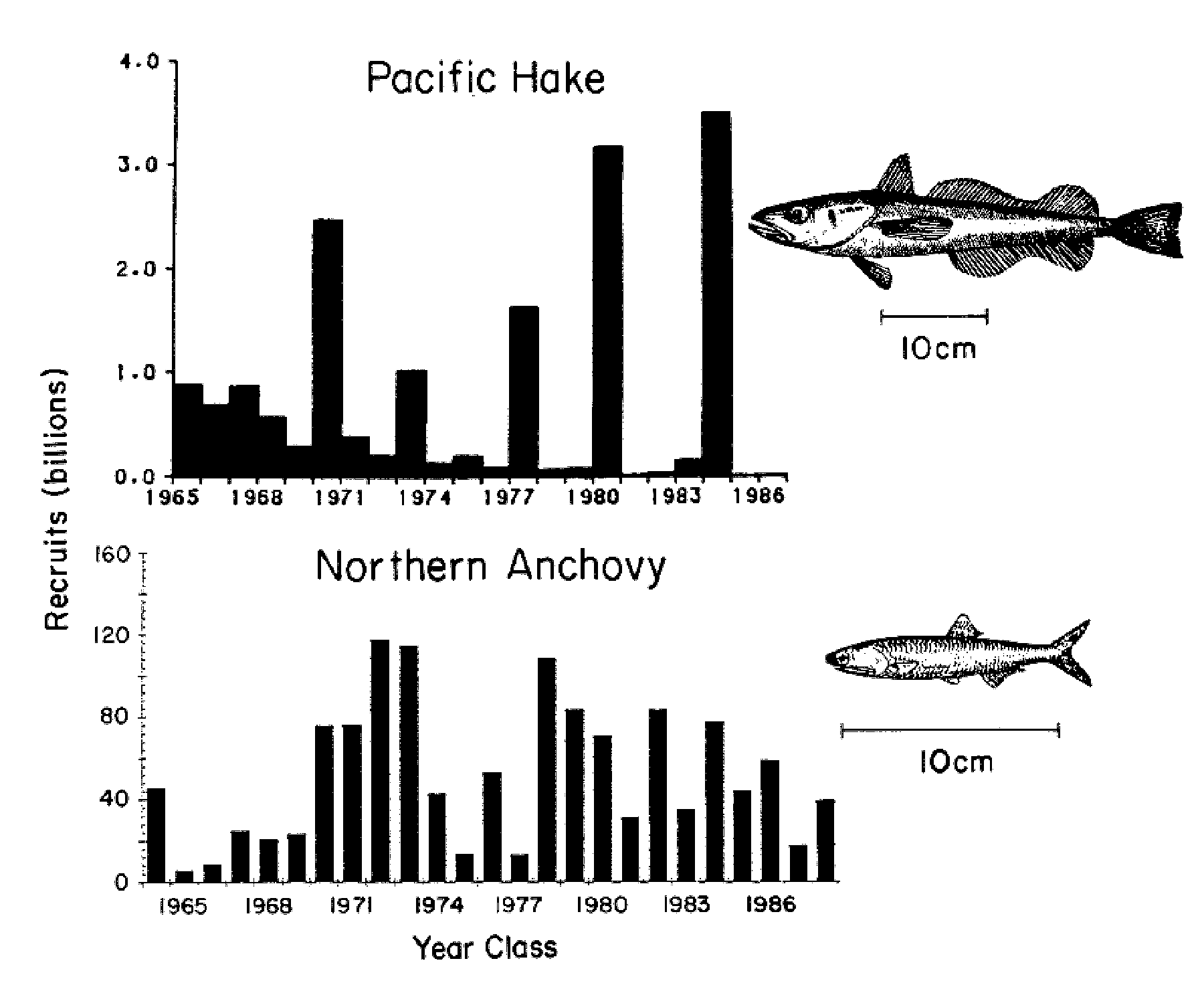
Figure 7. Comparative recruitment time series of Pacific hake
(Merluccius productus; from Hollowed and Bailey 1989) and northern
anchovy (Engraulis mordax; from Jacobson and Lo
1989).
Holozooplankton
Target species of zooplankton include representative copepods, euphausiids, and a salp or doliolid.
Some species are found in different regions of the CCS, exhibiting different life-history traits such as
overwintering strategies and vertical migration in different parts of their ranges. Examples include the
assemblage Calanus pacificus oceanicus, C. pacificus californicus, and Calanus marshallae. A
copepod genus whose �life history differs markedly from that of the genus Calanus, yet
is also widely distributed in the CCS is the cyclopoid copepod Oithona. Oithona might be considered a
"steady-state" genus in contrast to the "opportunistic" characteristics of Calanus. Eucalanus bungii and
E. californicus are also likely to offer interesting contrasts.
With respect to euphausiids, Euphausia pacifica is distributed throughout the CCS, from the Gulf of
Alaska to Baja California (Brinton 1962). It is the dominant species at many localities and makes an
excellent candidate for contrasting studies in different regions of the CCS. The northern
metapopulations of E. pacifica have shorter growing seasons and greater age and size at maturity than
the southern metapopulations. Other potential candidates include Thysanoessa spinifera and
congeners. Since euphausiids and copepods are the dominant prey for the target fish species identified
above, we expect to advance understanding of the coupling between physical processes, zooplankton
production, and fish recruitment.
Salps, and to a lesser extent doliolids, have extraordinarily rapid growth rates and colonizing abilities.
Historical evidence suggests that they may predominate in some regions of the CCS (Berner 1967). It
is also known that major ENSO events affect the total thaliacean (salp, doliolid, pyrosome) biomass
much more than the copepod and euphausiid biomass (Smith 1985). Both observations suggest that a
salp or doliolid species should also be a focus of study. Experiments should be designed expressly to
understand the contrasting processes that select for either thaliacean or crustacean dominance, and the
resulting consequences for the pelagic ecosystem.
Benthos
Among inhabitants of the hard-bottom benthos, the Dungeness crab and barnacles of the genera
Balanus or Cthamalus have the best historical data bases and are likely candidates for further study.
Evidence from some studies suggests that adult-adult interactions on the bottom may generally be more
important for controlling community structure in Region I (e.g., Paine 1974), while cross-shore
transport of pelagic larvae and migration of upwelling fronts may more significantly govern population
growth in Region II (Roughgarden et al. 1988).